The Response of \(\bf G\)-matrices to Random Genetic Drift
and a stochastic eco-evolutionary modelling framework
What is a \(\bf G\)-matrix? ![]()

- Multivariate trait vector: \({\bf z}=\left(\begin{smallmatrix} z_1 \\ \vdots \\ z_d \end{smallmatrix}\right)\)
- Trait decomposition: \({\bf z}={\bf g}+{\bf e}\)
- \(\bf g=\) genetic component
- \(\bf e=\) “environmental” component (error, everything else)
- Genetic covariance structure: \({\bf G}=\mathrm{Cov}({\bf g})=\left(\begin{smallmatrix} G_{11} & \cdots & G_{1d} \\ \vdots & \ddots & \vdots \\ G_{d1} & \cdots & G_{dd} \end{smallmatrix}\right)\)
- First two principle components:
![]()
- Heritable component of variation
- Predicts response to selection \(\Delta\bar{\bf z}={\bf G}\,\nabla_{\bar{\bf z}}\ln\bar W,\quad\bar W=\) mean fitness
- First two principle components:
What is random genetic drift?
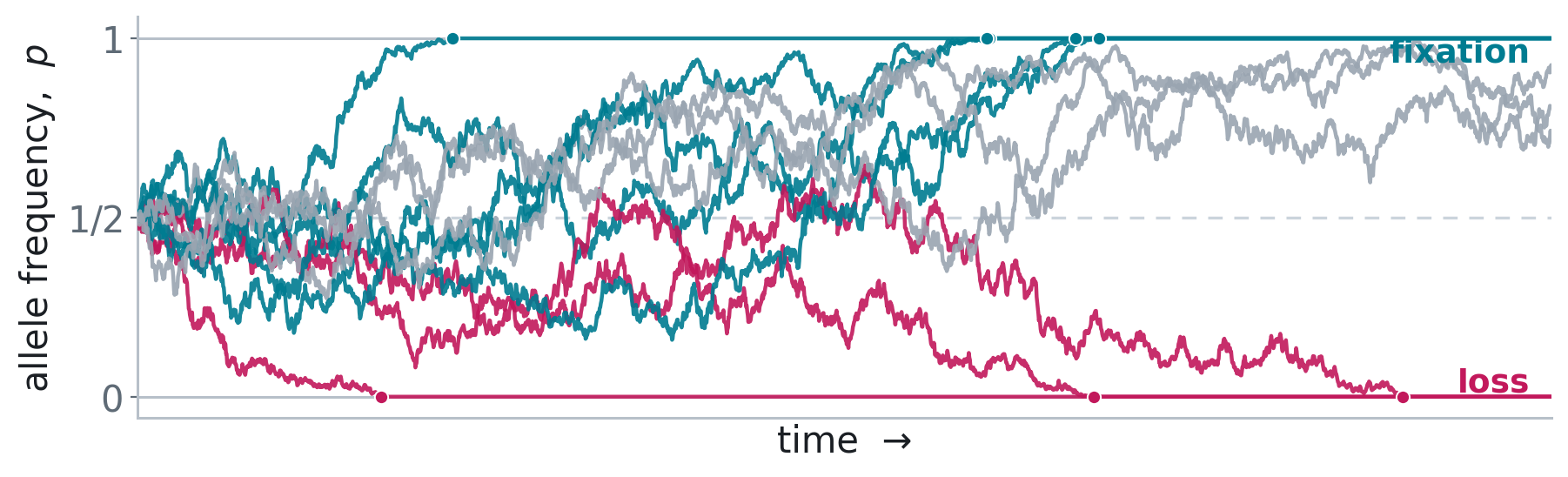
Response of \({\mathbb E}[{\bf G}]\) to drift
\(\tfrac{\mathrm d}{\mathrm dt}\mathbb E[{\bf G}]\approx-\tfrac{1}{N_e}\mathbb E[{\bf G}]\), (Lande, 1979, 1980)
\(\mathbb E[{\bf G}_t]\approx{\bf G}_0e^{-t/N_e}\)
Drift “shrinks” entries of \(\bf G\)-matrices
![]()

The conventional view
- Drift “shrinks” entries of \(\bf G\)-matrices
![]()
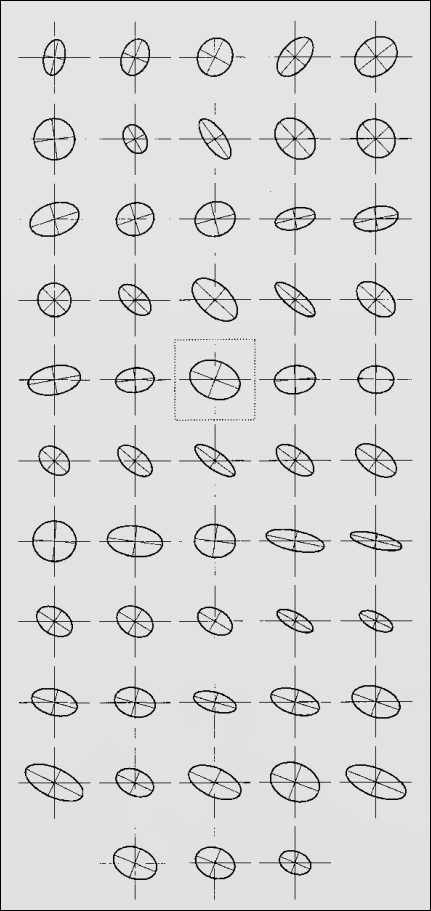
- No effect on genetic correlations between traits
- (btw, genetic correlations alter \(\bf G\)-matrix orientation)
![]()
- (btw, genetic correlations alter \(\bf G\)-matrix orientation)
- Differences in orientation ~ selection, mutation, migration (Roff 2000)
- Used as evidence for selection (Cano et al. 2004)

Empirical pushback and gap in theory
- Variation around \(\mathbb{E}[{\bf G}_t]\) (Phillips et al, 2001; Steppan et al, 2002)
- Still lacking stochastic neutral model (Mallard et al, 2024; Blomberg et al, 2025)
Why Take a Diffusion Limit?
Analytical tractability
Many models \(\rightarrow\) common diffusion limit
- Common leading-order structure
- Similar predictions across model class
- (can say a lot with a little?)
Not a universally better model
- Depends on scaling
- Features disappear in limit

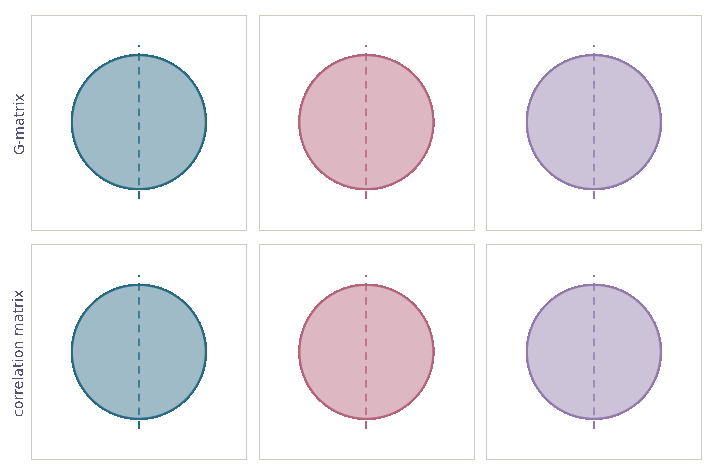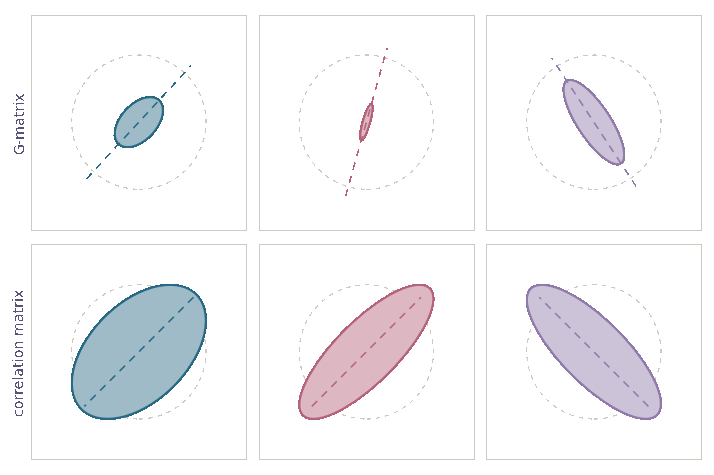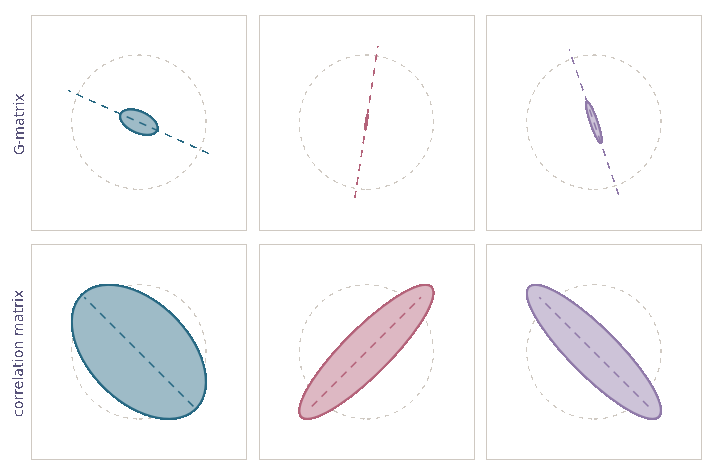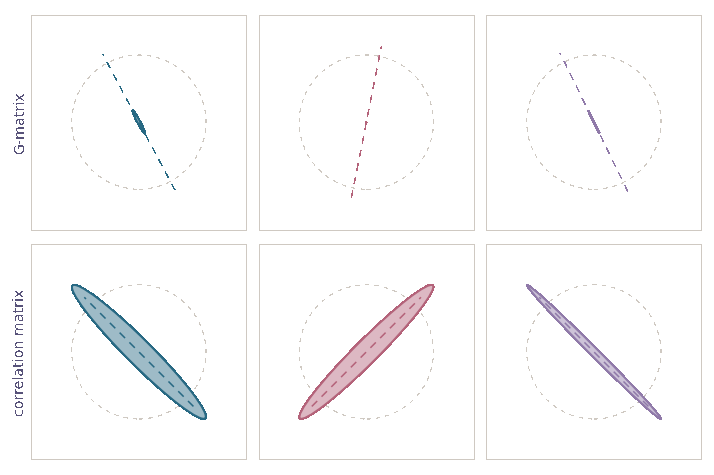
Genetic correlations tend towards ±1
\[\mathrm d\rho={\color{#c2185b}-\frac{1}{2N_e}\,\rho\,(1-\rho^2)}\,\mathrm dt+{\color{#5a4cc7}\sqrt{\frac{1}{N_e}\,(1-\rho^2)}\,\mathrm dB}\]
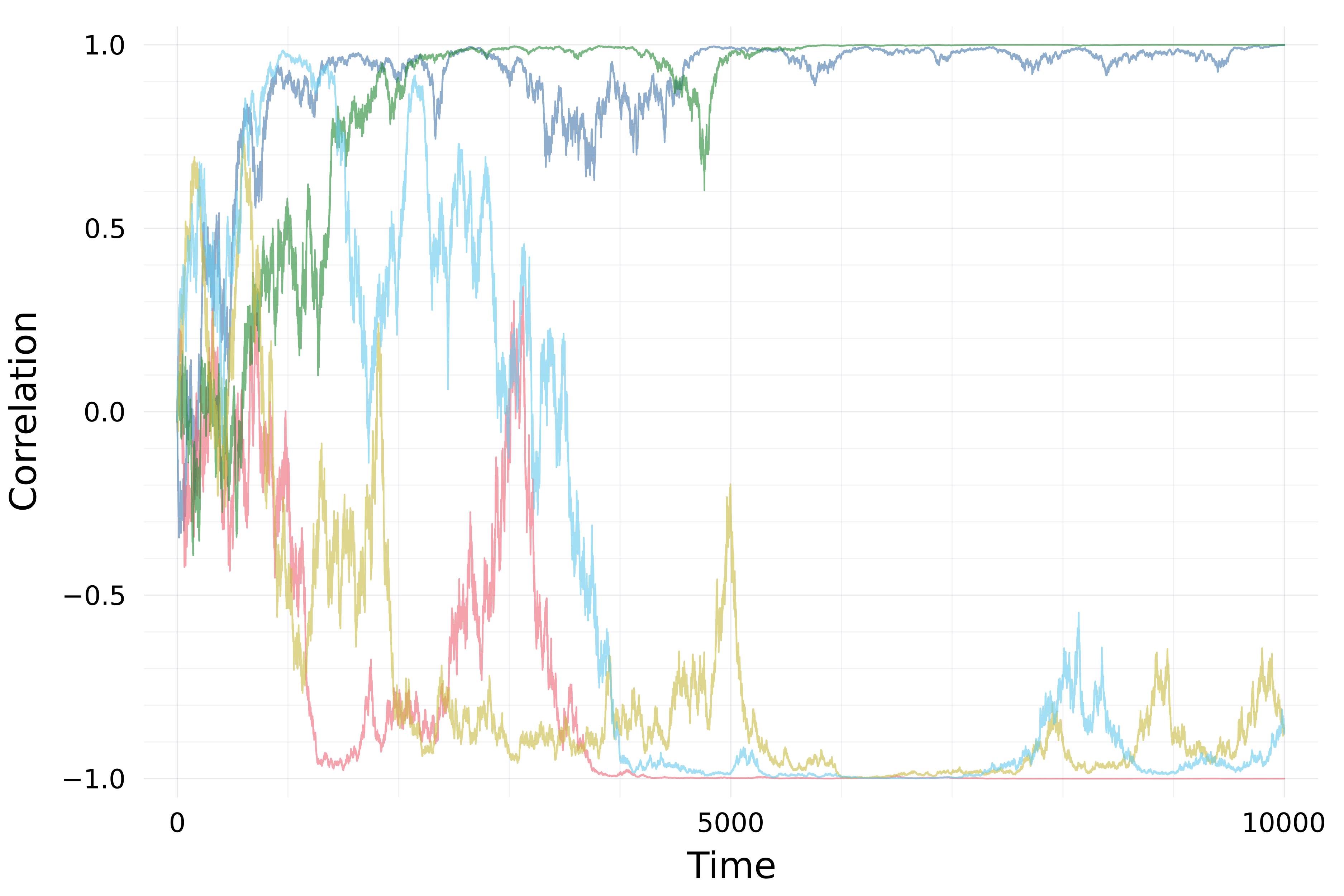
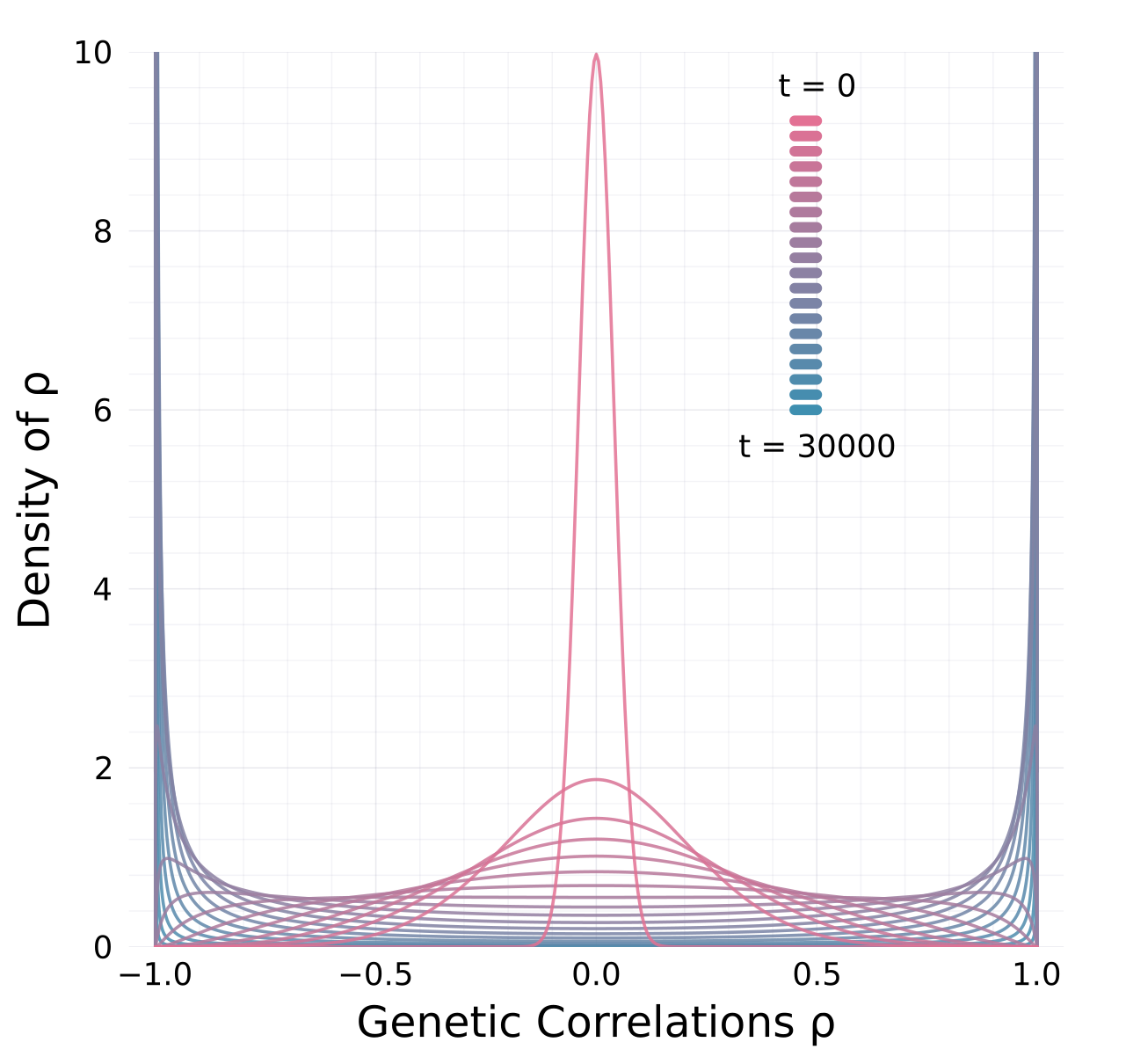
Revised view
- Drift alters orientation of \(\bf G\)-matrices
- while also eroding variation \({\mathbb E}[{\bf G}_t]\)
- Stability of \(\bf G\)-matrix structure due to
- mutation, migration, selection
- Essentially complete opposite of previous view (Roff 2000)
More in the paper…

Thanks & Questions
Steve Krone, Peter L. Ralph, Hinrich Schulenburg, Patrick C. Phillips, Arne Traulsen, Jonas Wickman, Brendan Bohannan
![]()



What about Recombination?
Drift alters \(\bf G\)-matrix orientation even with recombination
(Phillips et al. 2001)
(Mallard et al. 2023)